
 Índice
ÍndiceLa Química de la Vida ♦ Pabulo Henrique Rampelotto
Página 1 de 1.
![]()
 La Química de la Vida ♦ Pabulo Henrique Rampelotto
La Química de la Vida ♦ Pabulo Henrique Rampelotto
Manfenix Sáb Nov 30, 2013 2:36 pm

Actualmente existe un consenso en la comunidad científica de que los primeros seres vivos en nuestro planeta son el resultado de una larga evolución química, la cual precedió la evolución biológica. Esta hipótesis, inicialmente introducida por el bioquímico soviético A. I. Oparin, en 1924, e independientemente por el biólogo británico J. B. S. Haldane (en 1929, antes que el primer libro de Oparin fuese traducido para el inglés), postula que la vida se originó en la Tierra luego de una larga evolución de moléculas orgánicas simples hasta las más complejas, incluyendo macromoléculas de capacidad autorreplicativa.(1)
El experimento clásico de Stanley Miller fue la primera evidencia experimental de que la hipótesis de Oparin-Haldane podría estar correcta.(2) En este experimento, Miller usó una mezcla de los gases - metano (CH4), amoníaco (NH3) e hidrógeno (H2) - para simular la atmósfera primitiva y los electrodos para generar chispas que simularían los rayos y también serían la fuente de energía para las reacciones químicas. El análisis químico de la solución acuosa, algunas semanas después de iniciado el experimento, detectó la existencia de los aminoácidos glicina, α-alanina, ácido aspártico y α-aminoácido-n-butírico. Así, se demostró en el laboratorio la formación de compuestos orgánicos de interés biológico en condiciones similares a las de la atmósfera terrestre primitiva. Hoy, el experimento de Miller no es más considerado representativo para los procesos químicos que ocurrieron en la Tierra hace billones de años atrás, debido a que su modelo de la atmósfera primitiva no se encaja con el modelo actual. Nuevos estudios geoquímicos indican que la atmósfera terrestre nunca fue reductora (esto significa: formada por gases en sus formas más reducidas, como CH4, NH3 y H2), pero sí fue una atmósfera oxidante (CO2, CO, H2, N2) ó, en la mejor de las hipótesis, más o menos reductora, y en este tipo de atmósfera el rendimiento de aminoácidos producidos es muy bajo.(3)
Sin embargo, con este experimento, Miller inició el desarrollo de la investigación experimental sobre la evolución química y los orígenes de la vida.(4) Así, se puede decir que Miller es el fundador de lo que hoy se conoce como la química prebiótica, el estudio de las reacciones químicas que condujeron ó podrían haber conducido al surgimiento de la vida en nuestro planeta.
Con el desarrollo de poderosas técnicas de análisis química y molecular (por ejemplo, cromatografía gaseosa, espectroscopia de masa, cromatografía líquida de alta eficiencia y resonancia magnética nuclear) fue posible realizar estudios moleculares y quirales de los productos de las reacciones químicas.(5) Consecuentemente, hubo un gran avance en las investigaciones relacionadas a la química prebiótica. Gracias a estos trabajos, conocemos en detalles como muchas de las moléculas-llave de la vida pueden haber sido creadas en el mundo prebiótico. Los descubrimientos en las últimas décadas, sin embargo, han demostrado que los límites de la vida en la Tierra están lejos de ser definidos.(6) Para cualquier ambiente natural con condiciones extremas investigado, una variedad de microorganismos no tolera apenas, pero también requiere, estas condiciones para sobrevivir. Los microorganismos son encontrados a 6,7 km de profundidad en la corteza terrestre y a más de 10 km de profundidad en los océanos, sometidos a presiones de hasta 110 Mpa (1086 atm);(7) son encontrados también en ambientes extremamente ácidos (pH 0) hasta en ambientes extremamente básicos, pH 12,88 y en fuentes hidrotermales a la temperatura de 122 °C hasta en aguas heladas a −20 °C.(9) En adición a las rutas metabólicas de la fotosíntesis, microorganismos recientemente descubiertos poseen metabolismo basado en metano, azufre y hasta incluso hierro.(6)
Más allá de eso, es importante destacar que, a pesar de los recientes avances, estamos apenas comenzando a explorar y a caracterizar el mundo de los extremófilos. La mayor parte de los ambientes subterráneos y sub-oceánicos aún no fue estudiada y apenas un pequeño número de microorganismos ya fueron cultivados y descriptos.(10) Se estima que más de 90% de la flora microbiana permanece desconocida.(11) Por eso, los nuevos descubrimientos en el mundo microbiano pueden tener un gran impacto en nuestro conocimiento sobre la biosfera terrestre. En ese contexto, el reciente descubrimiento de Wolfe-Simon et al.(12) Tiene el potencial de reevaluar nuestro conocimiento de la biología. La autora y sus colaboradores publicaron el descubrimiento de una nueva bacteria que puede substituir arsénico (un elemento considerado altamente tóxico para los seres vivos) por fósforo en macromoléculas como los ácidos nucleicos y las proteínas. Por su naturaleza controversial, este trabajo viene recibiendo diversas críticas. Las principales hablan a respecto de la alta inestabilidad de los ésteres de arsénico en comparación a los ésteres de fosfato en el DNA y a la falta de evidencias directas de que el arsénico esté siendo realmente incorporado al DNA.(13) Algunos investigadores sugieren que la bacteria en cuestión está simplemente absorbiendo y aislando el arsénico en los vacuolos, mientras hace uso del fósforo.(14) Resultados más claros serán posibles cuando sea confirmada (ó no) la presencia de arsénico en el DNA ó RNA usando espectroscopia de masa. Si fuese correcto, este descubrimiento ampliaría nuestra definición de la vida como la conocemos.
En el laboratorio, el advenimiento de nuevas técnicas moleculares viene permitiendo la construcción de nuevas enzimas y otras macromoléculas.(15) Un genoma viral ya fue construido a partir de los oligonucleótidos.(16) En 2010, fue anunciada la creación de una célula bacteriana controlada por un genoma completamente artificial.(17) De esa forma, la emergente área científica de la biología sintética puede proveer ejemplos en el laboratorio de estructuras químicas alternativas capaces de soportar la catálisis y la evolución darwiniana.(18) con éstas técnicas revolucionarias tenemos el potencial de explorar si es posible construir la vida con nuevas fisiologías, incluyendo rutas metabólicas aún no utilizadas por los seres vivos.
Estos nuevos estudios y descubiertas, que desafían los paradigmas de la biología moderna, nos fuerzan a repensar cuestiones tan intrigantes como "¿qué es la vida?", "¿cuáles son sus límites?" y principalmente, "cuáles son sus características fundamentales?". En este artículo, estas cuestiones fueron estudiadas con el objetivo de ampliar el conocimiento sobre la naturaleza fundamental de los seres vivos, siendo ellos conocidos ó aún a ser descubiertos por la ciencia. En la primer parte, se discuten las dificultades de cómo definir la vida y sus implicaciones en la búsqueda por la vida extraterrestre. En la secuencia, se estudia si la posible química de formas alternativas de la vida y donde estas hipótesis pueden ser colocadas a prueba en nuestro sistema solar.
¿Qué es la vida?
Durante siglos los teólogos, filósofos y científicos han intentando encontrar una respuesta para esta cuestión.(19) Hay centenas de definiciones convencionales en la literatura, pero ninguna es capaz de alcanzar un consenso dentro de la comunidad científica. Existen definiciones termodinámicas, metabólicas, bioquímicas, genéticas y fisiológicas, para citar las más comunes.(20) Generalmente esas definiciones se resumen a meras listas de propiedades observables. Eso significa que aunque la caracterización de la vida sea posible, una definición precisa del concepto de vida aún está lejos de ser establecida. Cada teoría, hipótesis ó punto de vista adopta una definición, de acuerdo con sus propias premisas e intereses científicos.
Actualmente, el concepto más utilizado define a la vida como un "sistema químico auto-sustentable capaz de sufrir la evolución darwiniana".(21) Esa definición fue establecida en 1994 y desde entonces es adoptada por la NASA. Sin embargo, la capacidad de sufrir la evolución darwiniana, un proceso que resulta en mudanzas heredables en una población, no es suficiente para definir la vida, si consideramos que la capacidad de algunos minerales de reproducir los errores en su estructura cristalina sea equivalente a la evolución. De la misma forma, el concepto de un sistema químico autosustentable puede ser aplicado con alguna justificativa para otros procesos físico-químicos autosustentables, tales como los incendios forestales.
De modo general, las tradicionales definiciones de la vida, que se resumen a meras listas de propiedades observables, fallan en distinguir, de modo consistente, entre el mundo vivo y el no vivo. Lógicamente, esa distinción entre los dos no es difícil de ser hecha en nuestro planeta (pero, incluso para nuestro caso, algunos investigadores argumentan la existencia de una biosfera oculta, esto es, una biosfera que no somos capaces de detectar simplemente por ser diferente de aquella con la que estamos acostumbrados).(22) Pero una definición adecuada se vuelve relevante cuando salimos del contexto familiar de la Tierra y encontramos condiciones más exóticas y ambientes donde los fenómenos existen de modo completamente desconocido para nosotros. En esos casos, ¿cómo reconocer qué es vida y qué no lo es?
El único consenso que de hecho existe entre los científicos es que, a pesar de la gran biodiversidad, todos los seres vivos en nuestro planeta derivan de un ancestral común. Así, puede ser prematuro el intentar definir la vida basándose en un único ejemplo – la vida en la Tierra. Como expuesto por Cleland y Chyba,(23) las definiciones apenas nos dicen sobre el significado de las palabras en nuestra lengua, en lugar de decirnos sobre la naturaleza del mundo. Lo que realmente necesitamos es de una teoría general de los seres vivos, análoga a la teoría molecular, que nos permite dar una respuesta sin ambigüedad para la cuestión "¿qué es el agua?".(24) Antes de la teoría molecular, lo mejor que un científico podía hacer para caracterizar el agua era definirla en términos de sus propiedades sensibles, tales como el ser incolora, inodora e insípida. Cuando se comprendió la naturaleza molecular de la materia, fue posible identificar el agua de forma que toda la ambigüedad desapareció: el agua es H2O. Así, una respuesta precisa para la cuestión "¿qué es el agua?" fue posible apenas cuando fue colocada dentro de una teoría científica apropiada.
La falta de una definición de la vida, en algunos casos, puede ser problemática para las ciencias biológicas. Eso queda evidente, por ejemplo, en la intensa discusión en relación a los virus: ¿son ellos seres vivos ó no?; y más recientemente por la sorprendente hazaña de la biología sintética, como fue demostrado por la síntesis química de un genoma bacteriano completo y su incorporación en un microplasma.(17) De la misma forma, esa problemática se refleja en los debates generados por los avances recientes en la investigación biomédica que recae sobre el aborto, la eutanasia y los organismos transgénicos, para citar los más comunes. Más allá de eso, esa cuestión tiene implicaciones profundas en la búsqueda por la vida extraterrestre, como será discutido en la próxima sección.
Astrobiología y la búsqueda por la vida extraterrestre
De modo general, la astrobiología es una rama emergente de la ciencia que estudia la vida en el universo.(25) Ese amplio campo de investigación incluye el estudio del origen de la vida en la Tierra y en otras partes del universo, así como la búsqueda por vida extraterrestre.(26) Con esta afirmación, una pregunta nos viene rápidamente a la mente: "¿qué estamos buscando exactamente?". De esa forma, una respuesta adecuada para la cuestión es "¿qué es la vida?". Es fundamental para la astrobiología, debido a que el diseño de los experimentos de detección de la vida a ser realizados por telescopios y sondas espaciales, depende de suposiciones sobre el qué es la vida y cuáles observaciones se tendrán en cuenta como prueba para su detección.
Hasta el momento, nosotros apenas conocemos la vida en la Tierra. Por lo tanto, l búsqueda por seres extraterrestres ha sido la búsqueda por la vida como nosotros la conocemos, basada en un único ejemplo. Se cree que exoplanetas capaces de tener vida orbitan dentro de la llamada "zona habitable", la región alrededor de una estrella en la cual el agua se encuentra en estado líquido y donde la atmósfera puede contener dióxido de carbono, agua y nitrógeno.(27) Consecuentemente, los científicos han buscado por bio-huellas producidas por seres vivos extraterrestres en nuestro sistema solar (esto es, en meteoros, planetas como Marte y Venus y lunas como Europa, Titán y Encélado) que se asemejan a las células de organismos terrestres, moléculas derivadas del metabolismo celular, así como los aminoácidos y nucleótidos similares a aquellos encontrados en las proteínas y en el DNA.
La vida probablemente es el resultado de contingencias físicas y químicas presentes en el mundo donde ella surgió. En otros planetas, donde hay ambientes similares a los de la Tierra, es muy más probable que pueda existir vida como nosotros la conocemos, esto es, que utiliza agua como solvente y compuestos carbónicos como bloques constructores.(28) Sin embargo, aún no sabemos si realmente es lógico asumir ese pensamiento geocéntrico. La Tierra puede ser apenas uno de muchos modelos de planetas que pueden sustentar la vida. Basados en nuestro limitado conocimiento sobre el asunto, no se puede simplemente excluir la posibilidad de existir vida en ambientes muy diferentes al de la Tierra, donde un conjunto de compuestos químicos complejos, solventes y fuentes de energía se vuelven posibles a temperaturas, presiones y composiciones químicas diferentes de las de las condiciones terrestres.
De ese modo, las formas de vida que pueden haberse originado en otro lugar, aún dentro de nuestro propio sistema solar, pueden ser irreconocibles cuando son comparadas con la vida en la Tierra, no siendo posible detectarlas por telescopios y sondas espaciales destinadas a buscar biomoléculas terrestres ó sus productos.(29) Debemos reconocer que nuestro conocimiento sobre las características fundamentales de la vida está basado en nuestra comprensión de la biosfera durante las últimas fases de la historia de la Tierra.(30) Dado el hecho de que sólo conocemos un ejemplo de estructuras moleculares para la vida, es difícil imaginar como pueda ser la misma, en ambientes muy diferentes de los que encontramos en la Tierra. En las últimas décadas, sin embargo, los experimentos en el laboratorio y trabajos teóricos han venido indicando que la vida puede ser basada en estructuras moleculares substancialmente diferentes de aquellas que conocemos. A seguir, se discuten algunas de esas posibilidades con base en la literatura científica.
Posibles químicas alternativas para la vida
Aunque aún no haya una definición exacta sobre la vida, es posible identificar tres requisitos básicos para poder existir un ser vivo.(31) Cualquier forma de vida requiere la presencia de una gran cantidad y variedad de moléculas complejas para interaccionar con el medio externo y mantener su funcionamiento interno. De la misma forma, para mantener su alta complejidad química, la vida necesita de una fuente externa de energía y, finalmente, un medio líquido es necesario para mantener la concentración y, al mismo tiempo, el libre flujo de compuestos, permitiendo rápidas interacciones moleculares. Basados en esos tres requisitos básicos y obedeciendo a los principios físicos y químicos existentes, discutimos, a seguir, como la vida puede existir de un modo aún desconocido para nosotros.
Complejidad química
Una de las características fundamentales de la vida es su complejidad química, basada en moléculas poliméricas formadas por enlaces covalentes. El carbono parece ser el único elemento capaz de formar polímeros que rápidamente sufren alteraciones químicas bajo las condiciones físicas prevalecientes en la Tierra. Debido a sus propiedades químicas, el elemento carbono puede formar un número casi ilimitado de moléculas, siendo que algunas de ellas son comunes en el medio interestelar.(32) En consecuencia, se espera que cualquier forma de vida que pueda ser encontrada en otros planetas sea basada en el carbono. Sin embargo, de acuerdo con nuestro conocimiento actual de la física y de la química, ésta puede ser apenas una concepción geocéntrica.
Aunque una bioquímica basada en polímeros de cualquier átomo diferente del carbono generalmente reciba poca atención de la comunidad científica, estudios recientes sugieren que un organismo puede tener un metabolismo completamente independiente del carbono, esto es, un metabolismo cuyo esqueleto principal de las moléculas responsables por las reacciones químicas sea formado por otros elementos más allá del carbono.(33) En esos estudios, el silicio es el más citado por estar directamente abajo del carbono en la tabla periódica y, consecuentemente, ser el elemento más semejante químicamente. Algunas de las semejanzas y diferencias entre el silicio y el carbono que pueden llevar a alteraciones en las propiedades físico-químicas y biológicas de moléculas son discutidas por Bains y Tacke, Showell y Mills.(34) Como el carbono, el silicio puede formar cuatro enlaces, enlaces covalentes estables con él mismo y con otros elementos, más allá de formar compuestos estables tetra, penta y hexacoordenados con N, C y O.(35) De hecho, una gran variedad de estructuras vienen siendo formadas de tal química (Figura 1). Más allá de eso, la mayor reactividad del silicio comparada con la del carbono puede ser una ventaja en ambientes extremadamente fríos, muy comunes en el universo.
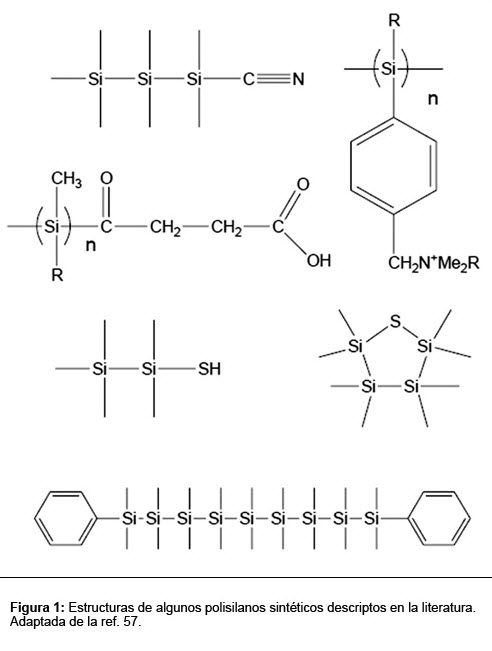
Lógicamente, es importante resaltar que, a pesar de algunas características semejantes, la química del silicio es más limitada que la del carbono. Sin embargo, las críticas en relación a los estudios con silicio, generalmente, son basadas en la improbabilidad de la vida basada en ese elemento bajo las condiciones ambientales terrestres. Es evidente que la substitución directa del carbono por silicio en la bioquímica terrestre produciría moléculas que serían casi inmediatamente hidrolizadas en el agua. Ciertos compuestos derivados del silicio, como silanos y muchos siloxanos, son espontáneamente inflamables a 0°C.(36) Sin embargo, estudios con los silanos indican que estos compuestos podrían disolverse en nitrógeno líquido, en lugar de en agua, en concentraciones suficientes para ser los precursores de la vida.(37) Es importante resaltar que los silanos son moléculas relativamente grandes, complejas y que pueden llevar a la producción de polímeros complejos.(38) Así, su flexibilidad estructural y química en medio no acuoso puede generar análogos funcionales a la bioquímica terrestre.
Los elementos azufre, nitrógeno y fósforo también son potenciales candidatos para formar el esqueleto principal de moléculas bioquímicas.(39) El azufre es capaz de formar largas cadenas moleculares, como el carbono. Algunas bacterias terrestres fueron descubiertas en ambientes ricos en azufre en lugar de oxígeno, reduciendo el azufre a sulfuro de hidrógeno.(40) El fósforo es similar al carbono en su habilidad de formar largas cadenas moleculares, que pueden llevar a la formación de macromoléculas complejas. Cuando es combinado con el nitrógeno, puede crearse una gran variedad de moléculas, incluyendo anillos.
Incluso elementos altamente tóxicos para los seres vivos, como el arsénico, pueden ser aprovechados por formas de vida bajo determinadas condiciones. En la Tierra, algunas algas marinas incorporan cantidades substanciales de arsénico en moléculas orgánicas complejas, como arseno-azúcares y arsenobetanoides.(41) Recientemente, fue descubierta una nueva bacteria que puede substituir arsénico por fósforo en macromoléculas, como los ácidos nucleicos y las proteínas.(14) Del mismo modo, muchas otras formas de vida microscópica utilizan arsénico para generar energía y facilitar su crecimiento.
Fuente de energía
Para mantener su alta complejidad química, la vida necesita de energía. Así, una fuente externa de energía es considerada fundamental para cualquier ser vivo. La energía proveniente del Sol es una fuente casi ilimitada para la vida en la Tierra, siendo muy importante para el desarrollo de la biosfera terrestre.
Con todo así, hay una gran cantidad de seres vivos que sobrevivieron usando otras fuentes de energía. Esos organismos obtienen energía a través de reacciones de óxido-reducción. En esos casos, los metabolismos más conocidos envuelven reacciones con elementos químicos como hidrógeno, hierro y azufre.(42) Sin embargo, muchas otras reacciones redox que generan energía para los seres vivos son conocidas y envuelven la oxidación y la reducción de elementos relativamente raros como el cobre, manganeso, arsénico y selenio.(43) Estos ejemplos demuestran la capacidad de la vida de usar diferentes formas de obtener la energía para su sobrevivencia. Se debe recordar que los seres vivos, de modo general, son capaces de detectar calor, presión y gravedad, así como los campos magnético y eléctrico.(44) Todos estos estímulos generan respuestas por la transducción de señales a través de la membrana celular, sea alterando la propagación de los iones que altera el potencial transmembrana, ó iniciando alteraciones metabólicas a través de sistemas de mensajero secundario. Eso indica como formas variadas de energía pueden afectar procesos biológicos. En este sentido, estudios teóricos han demostrado que otras fuentes de energía pueden ser tan eficientes como las energías luminosa y química (reacciones redox) presentes en la Tierra para sustentar seres vivos bajo un conjunto diferente de condiciones ambientales.(45)
En nuestro sistema solar varias fuentes de energía, además de la luz y las reacciones redox, están ampliamente disponibles y podrían ser utilizadas por los seres vivos.(46) La energía geotérmica es indicada por el vulcanismo, resultando en el calentamiento radiogénico en cuerpos planetarios como Venus y Marte. La presión es una fuente de energía en planetas con atmósfera espesa, como los gigantes gaseosos y Venus. La energía cinética es una fuerza presente en cualquier cuerpo planetario con fuerte convección, como los gigantes gaseosos y, posiblemente, Europa. La aceleración de las mareas es una fuente de energía comúnmente encontrada en el sistema solar, por ejemplo en Europa y Tritán. Los fuertes campos magnéticos y las radiaciones emanadas de Júpiter y Saturno afectan sus satélites. De la misma forma, la presión osmótica puede estar presente en las lunas con subocéanos líquidos, tales como Europa, Ganímedes y Titán.(46)
Lógicamente, eso no significa que tales fuentes de energía sean realmente usadas por seres hipotéticos, pero que apenas están presentes y por lo tanto, llenan uno de los requisitos básicos para la emergencia de la vida en un cuerpo planetario.
Medio líquido ó solvente
La vida, como la conocemos, consiste en interacciones químicas que ocurren en un medio líquido, una vez que las macromoléculas necesitan ser físicamente estables y, al mismo tiempo, presentar flexibilidad estructural y reactividad química. En la Tierra, el único líquido estable para los patrones físicos existentes es el agua, que también es compatible con gran parte de la química orgánica. Evidencias recientes indican que otros cuerpos planetarios en nuestro sistema solar pueden abrigar agua.(47) Tales evidencias llevaron a la NASA a adoptar el principio "siga el agua" en la búsqueda por vida extraterrestre. Así, el agua, como el carbono, son considerados esenciales para la vida.
Sin embargo, nuevos estudios vienen demostrando que varios otros solventes pueden sustentar reacciones bioquímicas (Tabla 1).(48) Entre ellos, el amoníaco es frecuentemente el solvente más citado.(49) El amoníaco, como el agua, disuelve diversos compuestos orgánicos. Muchas reacciones orgánicas preparadas en el laboratorio son hechas con este solvente. Más allá de esto, éste también es líquido en un amplio intervalo de temperaturas (Figura 2).
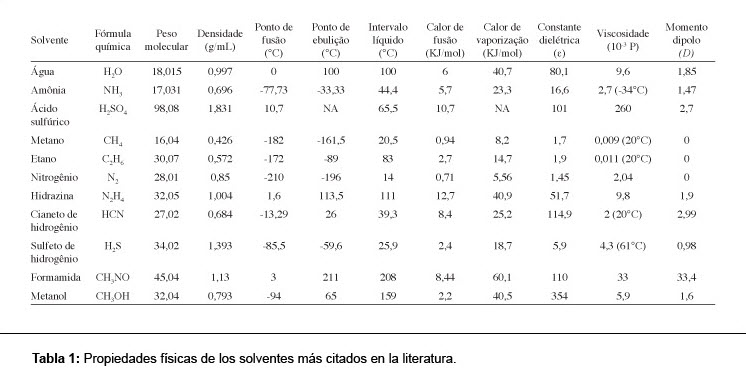
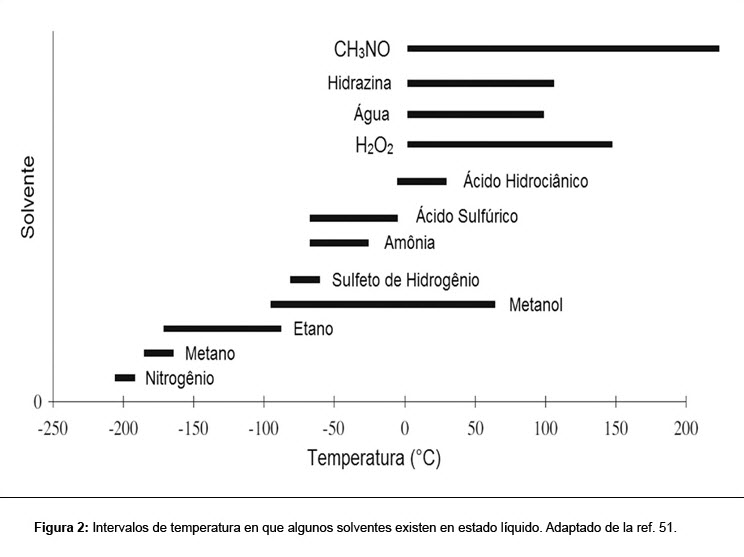
Las críticas en relación a los estudios con amoníaco resaltan que la formación de micelas por compuestos apolares en este solvente sería difícil. Sin embargo, la separación de fase hidrofóbica es posible en amoníaco en bajas temperaturas. Diferentes hidrocarburos se volvieron miscibles con amoníaco en diferentes temperaturas y presiones.(50) Así, la formación de las fases fóbica y fílica en amoníaco, análogas a las fases hidrofóbica y hidrofílica en agua, sería concebible en amoníaco, dependiendo de las condiciones de temperatura y presión escogidas. La agresividad del amoníaco como un nucleófilo implica que la bioquímica con base en ese solvente debería de ser diferente en comparación con la vida terrestre, como será discutido más adelante. Considerando que el amoníaco en su forma líquida puede ser abundante en el universo,(51) su potencial capacidad de sustentar vida extraterrestre es digna de consideración. El amoníaco no es el único solvente polar que puede ser considerado como una alternativa al agua. El ácido sulfúrico y la formamida son solventes que soportan reactividad química.(52) Más allá de eso, no hay necesidad de enfocar en solventes polares cuando se considera posibles hábitats para la vida. Los hidrocarburos no polares, como el metano y el etano, son mejores que el agua para el mantenimiento de reacciones químicas complejas, debido a que no destruyen hidrolíticamente las especies orgánicas inestables.(49) A pesar de su importancia para la química de la vida terrestre, en la mayoría de las veces, los químicos usan un solvente diferente del agua para ejecutar sus reacciones, pues ella propia es reactiva, presentando tanto un oxígeno nucleofílico como un hidrogeno ácido en concentraciones de 55 molares.
Es importante resaltar que los estudios experimentales envolviendo químicas alternativas para la vida son escasos y muchas de las ideas discutidas en ese área aún son especulativas. Con los recientes descubrimientos de posibles mundos habitables en nuestro sistema solar, este campo de investigación está comenzando a florecer. En la siguiente sección, se presentan algunas hipótesis de formas alternativas de vida que nuestro conocimiento actual permite discutir.
Formas alternativas de vida en nuestro sistema solar
Cuando se discute sobre la búsqueda por vida extraterrestre, una de las cuestiones más intrigantes que surge es "¿como tales formas exóticas de vida podrían ser?" ó "¿cuán semejantes ó diferentes de nosotros ellas serían?"
La vida es probablemente el resultado de contingencias físicas y químicas presentes en el mundo donde ella surge. Pero, aún en las condiciones existentes en la Tierra primitiva no son claramente comprendidas, lo que vuelve al origen de la vida terrestre un problema lejos de ser resuelto.(53) Más allá de eso, la historia de la vida en la Tierra muestra que la trayectoria evolutiva de un ser vivo no puede ser prevista. Las diversas formas inimaginables que surgieron durante el período Cambriano son un buen ejemplo de la variedad de formas que la vida puede tomar. De ese modo, las particularidades de la forma y la función de una historia evolutiva de la vida en otros mundos no pueden ser conocidas hasta ser encontradas. Mientras, a pesar de esa potencial diversidad, en el nivel molecular los mecanismos básicos orientan el desenvolvimiento de cualquier ser vivo. Así, con base a principios generales de la bioquímica es posible hacer previsiones sobre la naturaleza de formas exóticas de vida que pueden ser encontradas en el universo. A seguir, discutimos algunas de esas posibilidades en nuestro sistema solar, la única región del universo que podemos estudiar detalladamente en las próximas décadas.
Vida a base de amoníaco
El amoníaco, como fue revelado al explicar sus propiedades físicas, puede ser un buen solvente para la vida (Figura 3).(54) En realidad, las macromoléculas, como las proteínas, los aminoácidos y los ácidos nucleicos, contienen grupos funcionales OH y NH2 en varias combinaciones y proporciones en que el amoníaco puede fácilmente interaccionar. Sin embargo, una bioquímica con base en ese solvente debería ser diferente en comparación con la vida terrestre. Debido a que el oxígeno oxida y disocia las moléculas de amoníaco, la vida a base de amoníaco necesita de un ambiente sin la presencia del oxígeno.(55) En esos ambientes, el metabolismo anaeróbico sería la alternativa. Análogos de biomoléculas terrestres, en los cuales los átomos de oxígeno son substituidos por grupos NH, pueden producir una bioquímica igualmente viable (Figura 4). Reacciones de síntesis, como la de proteínas, a partir de aminoácidos a través de un enlace peptídico muestran semejanzas en la solución acuosa, amoníaca y mezclas de agua y amoníaco.(56)


Diversas líneas de evidencia sugieren que el océano interno de Titán contiene amoníaco mezclado con agua, bajo la forma de una capa líquida abajo de una corteza de hielo.(57) Más allá de eso, su ambiente libre de oxígeno proporciona una excelente oportunidad para encontrar seres vivos que utilizan amoníaco como solvente. La presencia de NH3 en el probable océano interno de Encélado también llamó la atención para la posibilidad de vida a base de amoníaco en este satélite.(58)
Vida a base de metano
Las evidencias recientes de lagos compuestos de hidrocarburos en la superficie de Titán, uno de los satélites de Saturno, han llevado a diversos científicos a estudiar la posibilidad de que esos líquidos sean un solvente, en el cual las formas de vida pueden desenvolverse.(59) Los estudios indican que estos lagos son compuestos de metano y etano, con trazas de nitrógeno disuelto.(60) Un ciclo de metano puede existir en Titán, semejante al ciclo hidrológico en la Tierra.(61) Así, esos reservatorios líquidos en la superficie proporcionan un ambiente donde moléculas prebióticas, formadas en la rica atmósfera de Titán, se pueden acumular y diferenciar químicamente.(62) Considerando la cantidad y extensión de los lagos de metano y etano, si los seres vivos florecen en esos ambientes líquidos y consumen compuestos producidos en la atmósfera, se espera que tales organismos tengan un efecto sobre el medio ambiente próximo a la superficie.
En este sentido, hace algunos años atrás, un estudio sugirió un tipo de vida a base de metano en Titán. De acuerdo con los investigadores, los micro-organismos podrían hacer reaccionar el hidrógeno con el material orgánico proveniente de la atmósfera (como acetileno ó etano) para obtener energía. El producto liberado, en este caso, sería metano. El estudio concluyó que los resultados de la sonda Huygens podrían indicar la presencia de tales formas de vida por la disminución anormal de acetileno y etano, así como de hidrógeno en la superficie de Titán.(63) Investigaciones recientemente divulgadas parecen corroborar esa hipótesis. Basado en análisis computacionales, uno de esos trabajos prevé un intenso flujo de hidrógeno de la superficie para el interior de Titán.(64) Otro estudio relató una disminución de acetileno en la superficie.(65) Más allá de eso, se sabe que la concentración del etano en la superficie de Titán es menor del esperado, mientras que el metano es encontrado en abundancia.(66) Aunque estos resultados no sean pruebas de que realmente haya vida en Titán, son relevantes. Aunque la existencia de vida en Titán sea un asunto en abierto, este satélite de Saturno es ampliamente considerado a ser un laboratorio natural de química prebiótica en nuestro sistema solar.
En consecuencia de estos resultados promisores, nuevos grupos de investigación se vienen formando alrededor del mundo con el objetivo de estudiar en detalles como los seres vivos podrían desenvolverse utilizando solventes como el metano.(67)
Vida a base de ácido sulfúrico
En los últimos años, el ácido sulfúrico ha sido sugerido como un posible solvente para la vida. Entre sus características, es líquido en un amplio límite de temperaturas (de 10 a 337 °C), posee constante dieléctrica y momento dipolo mayores que el agua y presenta alta viscosidad.(49) Debido a su capacidad de soportar la reactividad química, el ácido sulfúrico puede ser un solvente razonable, capaz de sustentar el metabolismo en ambientes no acuosos (Figura 4).
La atmósfera de Venus es el ambiente más apropiado en nuestro sistema solar donde esa forma exótica de vida puede florecer. Las nubes de Venus son compuestas principalmente de aerosoles de ácido sulfúrico y la presencia de agua es escasa.(68) La capa de nubes a 50 km arriba de la superficie podría proveer un ambiente propicio para la existencia de vida, con presiones semejantes a las encontradas en la Tierra, y temperaturas de 20 a 80 °C.(69) Según hipótesis recientes, formas de vida podrían haberse originado en un océano primordial cuando la superficie del planeta era más joven y fría;(70) A lo largo del tempo, cuando el planeta comenzó un proceso de calentamiento (actualmente, la temperatura en la superficie de Venus llega a 460 °C) las formas de vida adaptadas a desenvolverse en el ambiente atmosférico sobrevivieron.(71) Para protegerse de la gran cantidad de radiación UV recibida, éstos seres vivos podrían usar el compuesto ciclo-octa-azufre (S8) que es encontrado en abundancia en la capa baja de la atmósfera venusiana y no reacciona con el ácido sulfúrico.
Un proceso semejante se observa en la Tierra donde las bacterias púrpuras sulfurosas, las bacterias verdes sulfurosas y algunas especies de cianobacterias depositan gránulos de azufre en la superficie externa de la célula.(72) Esas formas de vida venusianas podrían ser fototróficas, usando sulfuro de hidrógeno (H2S), el cual sería oxidado para producir gránulos de azufre elemental.(73) En la Tierra, las bacterias púrpuras sulfurosas utilizan ese proceso anoxigénico como fuente de energía.(74)
Vida a base de silicio
El silicio puede formar largas cadenas de silanos, siliconas y silicatos.(75) Entre ellos, los silanos fueron considerados los compuestos más adecuados para sustentar la vida.(33) Sin embargo, una forma de vida basada en silicio tendría que ser completamente diferente de la vida como la conocemos en la Tierra. Los silanos entran en combustión espontáneamente en contacto con oxígeno, formando silicato e hidrógeno molecular. Así, una bioquímica con base en tales compuestos requiere un ambiente libre de oxígeno. La afinidad del silicio por el oxígeno es tan fuerte que, si el silicio fuese colocado en un medio acuoso, formaría un reservatorio de sílice, retirando el oxígeno del agua.(76) De esa forma, el agua no es un solvente compatible para los compuestos de silicio. El metano, etano, nitrógeno ó cualquier compuesto que contenga grupos metílicos son solventes más compatibles para un ser vivo a base de silicio (Figura 5).
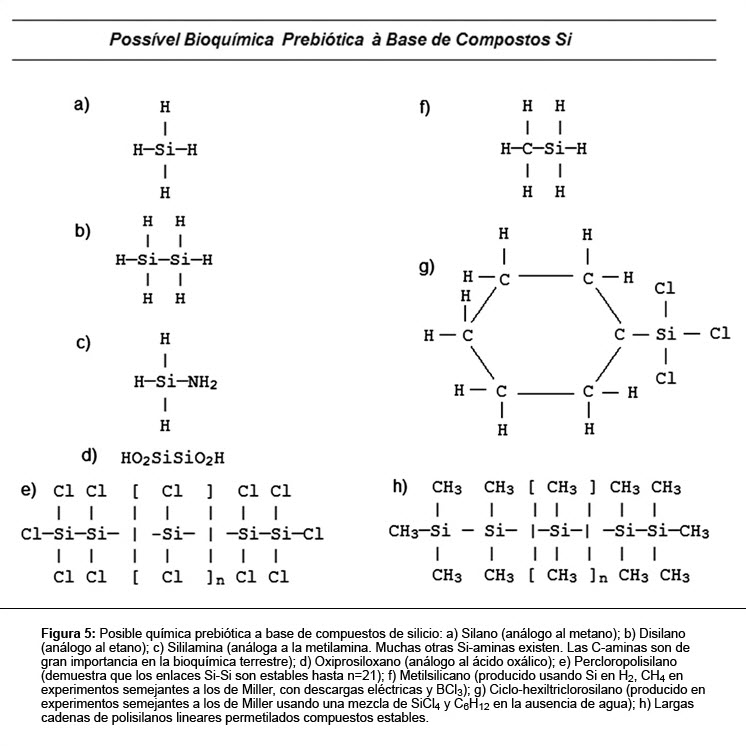
El enlace fuerte Si-O puede ser evitado y reproducido el escenario similar al carbono, si el oxígeno fuese substituido por azufre. Además, polímeros de silicio han sido obtenidos con nitrógeno en vez de oxígeno, donde el nitrógeno actúa como un donador de electrones.(77) En los ambientes pobres en hidrógeno, el mismo es frecuentemente substituido por un halógeno como cloro y son formadas largas cadenas lineares de silicio y cloruro.(78) Las grandes moléculas con base en el esqueleto NH-Si-Si, con halógenos como grupos laterales, podrían proveer la base para sistemas químicos complejos. Los silanos pueden formar macromoléculas flexibles con estructura variada.(79) Además de eso, los oligosilanos de hasta 26 enlaces Si-Si consecutivos pueden ser quirales, soportar una variedad de cadenas laterales funcionales y no funcionales, teniendo cadenas laterales alquilo, que son generalmente solubles en solventes apolares y autoagregados en estructuras anfifílicas en el agua, creando vesículas y micelas.(80)
Aunque la estabilidad de los silanos disminuye con el aumento de la cadena, si el hidrógeno fuese substituido por grupos orgánicos, se obtienen compuestos estables. Por ejemplo, polisilanos con pesos moleculares mayores que 100 ya fueron sintetizados.(81) Aunque los polisilanos no sean estables en las condiciones de temperatura y presión de la superficie de la Tierra, son suficientemente estables a bajas temperaturas, especialmente a altas presiones. Esos estudios sugieren que si la vida a base de silicio existe puede ser limitada a ambientes con pequeñas cantidades de oxígeno, escasez de agua, un solvente compatible (como metano ó nitrógeno) y temperaturas extremadamente bajas.
El Titán es un excelente destino en nuestro sistema solar para investigar esa posibilidad, pues satisface todos los criterios arriba descriptos.(82) Aunque se considere que la abundancia de compuestos carbónicos en Titán pueda competir con los compuestos de silicio como bloques constructores de la vida, el silicio puede ser una ventaja en ambientes extremadamente fríos, debido a su alta reactividad.
Tritón, la mayor luna de Neptuno, es otra posibilidad donde la vida a base de silicio puede ser estudiada. Este satélite es geológicamente activo, presentando actividad volcánica, más allá de poseer una tenue atmósfera y, en especial, un amplio reservatorio de nitrógeno líquido fluyendo abajo de la superficie, de la misma forma que la lava fluye abajo de la superficie terrestre.(83) Los estudios han demostrado que los silanos pueden disolverse en nitrógeno líquido en concentraciones suficientes para ser precursores de la vida. Así, Tritón posee los requisitos básicos para ser considerado un objetivo en potencial para estudios astrobiológicos.(84)
Consideraciones Finales
Nuestro amplio conocimiento de la química terrestre nos lleva a concentrar la atención en la química del agua y del carbono, compuestos en que la vida terrestre está basada y - como sabemos - la química orgánica surgió en el siglo XIX, con base al aislamiento de compuestos de la naturaleza. Esta tendencia para retratar la vida llevando en cuenta las características específicas de la vida de la Tierra es la base para lo que llamamos el "principio geocéntrico", discutido por Rampelotto.(29) Nuestra tendencia natural al geocentrismo requiere que se haga un gran esfuerzo para ampliar la percepción sobre las características fundamentales de la vida. Como sólo conocemos un ejemplo de vida, aún no sabemos todas sus potencialidades. Así, no podemos descartar la posibilidad de que las características más importantes de la vida aún no hayan sido descubiertas. Las características tradicionalmente apuntadas como esenciales a los seres vivos, pueden ser apenas consecuencias de las propiedades más fundamentales, pero aún desconocidas. En este sentido, los estudios aquí discutidos, exploran posibilidades alternativas de la vida, son fundamentales para la formulación de una teoría general de los sistemas vivos que, por su vez, puede guiarnos en dirección a una respuesta para la cuestión "¿qué es vida?".
Además de eso, principios básicos de la química nos advierten contra el pensamiento geocéntrico en relación a la vida. A través de los ejemplos discutidos, es posible imaginar reacciones químicas que puedan sustentar a los seres vivos envolviendo compuestos no carbónicos, ocurriendo en otros solventes más allá del agua, lo que envuelven reacciones de óxido-reducción, sin oxígeno molecular. Estos estudios sugieren que, si seres vivos se originaron de forma independiente en otros mundos que presentan ambientes geoquímicos diferentes, probablemente éstos habrían seleccionado diferentes solventes y bio-polímeros para el metabolismo. Así, en vez de buscar por bio-huellas específicas que surgieron recientemente en la historia de la Tierra (como, por ejemplo, oxígeno y ozono), las futuras misiones espaciales deben concentrarse en la búsqueda de vida basada en sus características fundamentales, lo que significa buscar por las características de la vida, independientemente del material que la constituye. Al asumir esta visión no geocéntrica, estaremos aumentando significativamente las probabilidades de reconocer un ser vivo, si nos encontramos con él, en algún lugar del universo.
Contacto
e-mail: [Tienes que estar registrado y conectado para ver este vínculo]
Autor
Pabulo Henrique Rampelotto - Centro Interdisciplinar de Pesquisas en Biotecnologia, Universidade Federal do Pampa, CP 1847, 97300-000 São Gabriel - RS, Brasil.
Referencias
1. Oparin, A. I.; The Origin of Life, Moscow Worker Publisher: Moscow, 1924 (em Russo); Haldane, J. B. S.; Rationalist Annual 1929, 3, 1.
2. Miller, S. L.; Science 1953, 117, 528.
3. Selsis, F. en Astrobiology: Future Perspectives; Ehrenfreund, P.; Irvine, W. M.; Owen, T.; Becker, L.; Blank, J.; Brucato, J. R.; Colangeli, L.; Derenne, S.; Dutrey, A.; Despois, D.; Lazcano, A.; Robert, F., eds.; Klu- wer Academic Publishers: New York, 2005; Heinrich, M. N.; Khare, B. N.; McKay, C. P.; Icarus 2007, 191, 765; Kasting, J. F. en Prebiotic Evolution and Astrobiology; Wong, J. T. F.; Lascano, A., eds.; Landes Bioscience: Austin, 2009.
4. Zaia, D. A. M.; Quim. Nova 2003, 26, 260.
5. Wong, J. T. F.; Lascano, A.; Prebiotic Evolution and Astrobiology, Lan- des Bioscience: Austin, 2009; Rauchfuss, H.; Chemical Evolution and the Origin of Life, Springer-Verlag: Berlin, 2008.
6. Rampellotto, P. H.; Rosa, M. B.; Schuch, A. P.; Schuch, N. J.; Pinheiro, D. K.; Munakata, N.; Astrobiology 2007, 7, 528; Rampelotto, P. H.; Sustainability 2010, 2, 1602.
7. Pikuta, E. V.; Hoover, R. B.; Crit. Rev. Microbiol. 2007, 33, 183.
8. Rothschild, L. J.; Mancinelli, R. L.; Nature 2001, 409, 1092.
9. Cavicchioli, R.; Astrobiology 2002, 2, 281.
10. Rainey, F.; Oren, A.; Extremophiles, Academic Press: New York, 2006.
11. Epstein, S. S.; Uncultivated Microorganisms, Springer: New York, 2009.
12. Wolfe-Simon, F.; Blum, J. S.; Kulp, T. R.; Gordon, G. W.; Hoeft, S. E.; Pett-Ridge, J.; Stolz, J. F.; Webb, S. M.; Weber, P. K.; Davies, P. C. W.; Anbar, A. D.; Oremland, R. S.; Science 2011, 32, 1163.
13. Schwartz, A. W.; Orig. Life Evol. Biosph. 2011, 41, 1.; Tawfik, D. S.; Viola, R. E.; Biochemistry 2011, 50, 1128.
14. Katsnelson, A.; Nature 2010, 468, 741.
15. Rothschild, L. J.; BioEssays 2010, 32, 304.
16. Smith, H. O.; Hutchison III, C. A.; Pfannkoch, C.; Venter, J. C.; Proc. Natl. Acad. Sci. U.S.A. 2003, 100, 15440.
17. Gibson, D. G.; Glass, J. I.; Lartigue, C.; Noskov, V. N.; Chuang, R. Y.; Algire, M. A.; Benders, G. A.; Montague, M. G.; Ma, L.; Moodie, M. M.; Merryman, C.; Vashee, S.; Krishnakumar, R.; Assad-Garcia, N.; Andrews-Pfannkoch, C.; Denisova, E. A.; Young, L.; Qi, Z. Q.; Segall- Shapiro, T. H.; Calvey, C. H.; Parmar, P. P.; Hutchison, C. A.; Smith, H. O.; Venter, J. C.; Science 2010, 329, 52.
18. Morange, M.; EMBO 2009, 10, 50.
19. Tsokolov, S. A.; Astrobiology 2009, 9, 401.
20. Pályi, G.; Zucchi, C.; Caglioti. L.; Fundamentals of Life, Elsevier: New York, 2002; Popa, R.; Between Necessity and Probability: Searching for the Definition and Origin of Life, Springer: Berlin, 2004.
21. Joyce, G. F. en Origins of life: The Central Concepts; Deamer, D. W.; Fleischaker, G. R., eds.; Jones and Bartlett: Boston, 1994.
22. Cleland, C. E.; Stud. Hist. Phil. Biol. Biomed. Sci. 2007, 38, 847; Davies, P. C. W.; Phil. Trans. R. Soc. A 2011, 369, 624.
23. Cleland, C. E.; Chyba C. F.; Orig. Life Evol. Biosph. 2002, 32, 387.
24. Chyba, C. F.; Hand, K. P.; Annu. Rev. Astron. Astrophys. 2005, 43, 1.
25. Plaxco, K. W.; Gross. M.; Astrobiology: A Brief Iintroduction, The Johns Hopkins University Press: Baltimore, 2006; Ehrenfreund, P.; Irvine, W. M.; Owen, T.; Becker, L.; Blank, J.; Brucato, J. R.; Colangeli, L.; De- renne, S.; Dutrey, A.; Despois, D.; Lazcano, A.; Robert, F.; Astrobiology: Future Perspectives, Kluwer Academic Publishers: New York, 2005.
26. Des Marais, D. J.; Nuth, J. A.; Allamandola, L. J.; Boss, A. P.; Farmer, J. D.; Hoehler, T. M.; Jakosky, B. M.; Meadows, V. S.; Pohorille, A.; Runnegar, B.; Spormann, A. M.; Astrobiology 2008, 8, 715; Bennett, J.; Beyound UFOs: The Search for Extraterrestrial Life, Princeton Univer- sity Press: Princeton, 2008; Horneck, G.; Rettberg, P.; Complete Course in Astrobiology, Wiley-VCH: Darmstadt, 2007.
27. Rekola, R. T. F.; Planet. Space Sci. 2009, 57, 430; Fridlund, M.; Eiroa, C.; Henning, T.; Herbst, T.; Lammer, H.; Léger, A.; Liseau, R.; Paresce, F.; Penny, A.; Quirrenbach, A.; Röttgering, H.; Selsis, F.; White, G. J.; Absil, O.; Defrère, D.; Hanot, C.; Stam, D.; Schneider, J.; Tinetti, G.; Karlsson, A.; Gondoin, P.; den Hartog, R.; D'Arcio, L.; Stankov, A. M.; Kilter, M.; Erd, C.; Beichman, C.; Coulter, D.; Danchi, W.; Devirian, M.; Johnston, K. J.; Lawson, P.; Lay, O. P.; Lunine, J.; Kaltenegger, L.; Astrobiology 2010, 10, 5.
28. Brack, A.; Horneck, G.; Cockell, C. S.; Bérces, A.; Belisheva, N. K.; Eiroa, C.; Henning, T.; Herbst, T.; Kaltenegger, L.; Léger, A.; Liseau, R.; Lammer, H.; Selsis, F.; Beichman, C.; Danchi, W.; Fridlund, M.; Lunine, J.; Paresce, F.; Penny, A.; Quirrenbach, A.; Röttgering, H.; Schneider, J.; Stam, D.; Tinetti, G.; White, G. J.; Astrobiology 2010, 10, 69.
29. Rampelotto, P. H. en Abiogenesis, Origins and the Search for Life; Russell, M., ed.; Cosmology Science Publishers: Cardiff, 2010.
30. Pilcher, C. B.; Astrobiology 2003, 3, 471.
31. Irwin, L. N.; Schulze-Makuch, D.; Astrobiology 2001, 1, 143.
32. Thaddeus, P.; Phil. Trans. Roy. Soc. B 2006, 361, 1681.
33. Bains, W.; Astrobiology 2004, 4, 137; Baross, J. A.; Benner, S. A.; Cody, G. D.; Copley, S. D.; Pace, N. R.; Scott, J. H.; Shapiro, R.; Sogin, M. L.; Stein, J. L.; Summons, R.; Szostak, J. W.; The Limits of Organic Life in Planetary Systems, National Academic Press: Washington, 2007.
34. Bains, W.; Tacke, R.; Curr. Opin. Drug Discov. Devel. 2003, 6, 526; Showell, G. A.; Mills, J. S.; Drug Discov. Today 2003, 8, 551.
35. Brook, M. A.; Silicon in Organic, Organometallic and Polymer Chem- istry, John Wiley: Toronto, 2000.
36. Koda, S.; Prog. Energy Combust. Sci. 1992, 18, 513.
37. Murugavel, R.; Chandresekhar, V.; Voight, A.; Roesky, H. W.; Schmidt, H. G.; Noltemeyer, M.; Organometallics 1995, 14, 5298; Lickiss, P. en The Chemistry of Organic Silicon Compounds; Rappoport, Z.; Apeloig, Y., eds.; John Wiley and Sons: New York, 2001.
38. Lickiss, P.; Adv. Inorg. Chem. 1995, 42, 174.
39. Ward, P.; Life las We Do Not Know It: The NASA Search for (and Syn- thesis of) Alien Life, Viking Adult: New York, 2005.
40. So, C. M.; Young, L. Y.; Appl. Environ. Microbiol. 1999, 65, 2969; Haouari, O.; Fardeau, M. L.; Casalot, L.; Tholozan, J. L.; Hamdi, M.; Ollivier, B.; Int. J. Syst. Evol. Microbiol. 2006, 56, 2909.
41. Barra, C. M.; Santelli, R. E.; de la Guardia, M.; Quim. Nova 2000, 23, 58.
42. Maier, R. M.; Pepper, I. L.; Gerba, C. P.; Environmental Microbiology, Academic Press: San Diego, 2000.
43. Madigan, M. T.; Martinko, J. M.; Stahl, D.; Clark, D. P.; Brock Biol- ogy of Microorganisms, 2nd ed., Benjamin Cummings: San Francisco, 2010.
44. Blakemore, R. P.; Frankel, R. B.; Sci. Am. 1981, 245, 42; Braucker, R.; Cogoli, A.; Hemmersbach, R. en Astrobiology: the Quests for the Conditions of Life; Horneck, G.; Baumstark-Khan, C., eds.; Springer: Berlin, 2002; Viswanath, V.; Story, G. M.; Peier, A. M.; Petrus, M. J.; Lee, V. M.; Hwang, S. W.; Patapoutian, A.; Jegla, T.; Nature 2003, 423, 822; Wilmer, P.; Stone, G.; Johnston, I.; Environmental Physiology of Animals, Blackwell: Oxford, 2000.
45. Muller, A. W. J.; Astrobiology 2003, 3, 555; Gusev, V. A.; Schulze- Makuch, D.; Naturwissenschaften 2005, 92, 115; Irwin, L. N.; Schulze- Makuch, D.; Cosmic Biology, Springer: New York, 2011.
46. Schulze-Makuch, D.; Irwin, L. N.; Life in the Universe: Expectations and Constraints, 2nd ed., Springer: Berlin, 2008.
47. Encrenaz, T.; Annu. Rev. Astron. Astrophys. 2008, 46, 57.
48. Baross, J. A.; Benner, S. A.; Cody, G. D.; Copley, S. D.; Pace, N. R.; Scott, J. H.; Shapiro, R.; Sogin, M. L.; Stein, J. L.; Summons, R.; Szostak, J. W.; The Limits of Organic Life in Planetary Systems, Na- tional Academic Press: Washington, 2007.
49. Schulze-Makuch, D.; Irwin, L. N.; Naturwissenschaften 2006, 93, 155.
50. Brunner, E.; J. Chem. Thermodyn. 1988, 20, 1397; Barajas-Fernándeza, J.; Alvarado, J. F. J.; González-Alatorre, G.; Estrada-Baltazar, A.; González-García, R.; Fluid Phase Equilib. 2009, 279, 92.
51. Lewis, J. S.; Physics and Chemistry of the Solar System, 2nd ed., Else- vier: London, 2004.
52. Naganuma, T.; Sekine, Y.; J. Cosmol. 2010, 5, 905.
53. Zaia, D. A. M.; Zaia, C. T. B. V.; Quim. Nova 2008, 31, 1599; Rampe- lotto, P. H.; J. Cosmol. 2009, 1, 86.
54. Fortes, A. D.; Icarus 2000, 146, 444; Neish, C. D.; Somogyi, A.; Lunine, J. I.; Smith, M. A.; Icarus 2009, 201, 412.
55. Benner, S. A.; Ricardo, A.; Carrigan, M. A.; Curr. Opin. Chem. Biol. 2004, 8, 672.
56. Firsoff, V. A.; Life Beyond the Earth: A Study in Exobiology, Basic Books: New York, 1963; Raulin, F.; Bruston, P.; Paillous, P.; Sternberg, R.; Adv. Space Res. 1995, 15, 321.
57. Tobie, G.; Grasset, O.; Lunine, J. I.; Mocquet, J.; Sotin, C.; Icarus 2005, 175, 496.
58. Waite, Jr.; J. H.; Lewis, W. S.; Magee, B. A.; Lunine, J. I.; McKinnon, W. B.; Glein, C. R.; Mousis, O.; Young, D. T.; Brockwell, T.; Westlake, J.; Nguyen, M. J.; Teolis, B. D.; Niemann, H. B.; McNutt Jr.; R. L., Perry; M.; Ip, W. H.; Nature 2009, 460, 487.
59. Rampelotto, P. H. en The Biological Big Bang: Panspermia and the Origins of Life; Wickramasinghe, C., ed.; Cosmology Science Publishers: Cardiff, 2010; Rampelotto, P. H.; 38th COSPAR Scientific Assembly Abstracts Book, Bremen, Germany, 2010.
60. Stofan, E. R., Lunine, J. I.; Elachi, C.; Nature 2007, 445,61; Lunine, J. I.; Lorenz, R. D.; Annu. Rev. Earth Planet. Sci. 2009, 37, 299.
61. Lunine, J. I.; Atreya, S. K.; Nature Geosci. 2008, 1, 160; Tokano, T.; McKay, C. P.; Neubauer, F. M.; Atreya, S. K.; Ferri, F.; Fulchignoni, M.; Niemann, H. B.; Nature 2006, 442, 432.
62. Raulin, F.; Nature 2008, 454, 587.
63. McKay, C. P.; Smith, H. D.; Icarus 2005, 178, 274.
64. Strobel, D. F.; Icarus 2010, 208, 878.
65. Clark, R. N.; Curchin, J. M.; Barnes, J. W.; Jaumann, R.; Soderblom, L.; Cruikshank, D. P.; Brown, R. H.; Rodriguez, S.; Lunine, J.; Stephan, K.; Hoefen, T. M.; Le Mouélic, S.; Sotin, C.; Baines, K. H.; Buratti, B. J.; Nicholson, P. D.; J. Geophys. Res. 2010, 115, E10005.
66. Lorenz, R. D.; Mitchell, K. L.; Kirk, R. L.; Hayes, A. G.; Aharonson, O.; Zebker, H. A.; Paillou, P.; Radebaugh, J.; Lunine, J. I.; Janssen, M. A.; Wall, S. D.; Lopes, R. M.; Stiles, B.; Ostro, S.; Mitri, G.; Stofan, E. R.; Geophys. Res. Lett. 2008, 35, L02206.
67. Norman, L. H.; Fortes, A. D.; A&G 2011, 52, 39; Leitner, J. J.; Schulze- Makuch, D.; Firneis, M. G.; Hitzenberger, R.; Neubauer, D.; 10th European Astrobiology Network Association Science Congress Abstracts Book, Pushchino, Russia, 2010.
68. Markiewicz, W. J.; Titov, D. V.; Limaye, S. S.; Keller, H. U.; Ignatiev, N.; Jaumann, R.; Thomas, N.; Michalik, H.; Moiss, R.; Russo, P.; Nature 2007, 450, 633.
69. Svedhem, H.; Titov, D. V.; Taylor, F. W.; Witasse, O.; Nature 2007, 450, 629.
70. Schulze-Makuch, D.; Grinspoon, D. H.; Abbas, O.; Irwin, L. N.; Bullock, M.; Astrobiology 2004, 4, 11.
71. Cockell, C. S.; Planet. Space Sci. 1999, 47, 1487.
72. Tortora, G.; Funke, B.; Case, C.; Microbiology: An Introduction, 5th ed., Addison Wesley Longman: San Francisco, 2001.
73. Schulze-Makuch, D.; Irwin, L. N.; Astrobiology 2002, 2, 197.
74. Herbert, R. A.; Gall, A.; Maoka, T.; Cogdell, R. J.; Robert, B.; Takaichi, S.; Schwabe, S.; Photosynth. Res. 2008, 95, 261.
75. Jutzi, P.; Schubert, U.; Silicon Chemistry: From the Atom to Extended Systems, Wiley-VCH: New York, 2003.
76. Le Grand, A. P.; The Surface Properties of Silicas, John Wiley and Sons: New York, 1998.
77. Murugavel, R.; Chandresekhar, V.; Voight, A.; Roesky, H. W.; Schmidt, H. G.; Noltemeyer, M.; Organometallics 1995, 14, 5298.
78. Tang, H.; Li, J.; Qin, J.; React. Funct. Polym. 2001, 48, 193.
79. Auner, N.; Weis, J.; Organosilicon Chemistry III: From Molecules to A Química da Vida como nós não conhecemos 1627Vol. 35, No. 8 Materials, Wiley-VCH: Weinheim, 1998; Fan, H.; Chen, Z.; Brinker, C. J.; Clawson, J.; Alam, T.; J. Am. Chem. Soc. 2005, 127, 13746; Mu- zafarov, A.; Silicon Polymers, Springer-Verlag: Berlin, 2011.
80. Sharma, H. K.; Pannell, K. H.; Chem. Rev. 1995, 95, 1351; Hengge, E.; Janoschek, R.; Chem. Rev. 1995, 95, 1495.
81. Rappoport, Z.; Apeloig, Y.; The Chemistry of Organic Silicon Com- pounds, John Wiley and Sons: New York, 2001.
82. Tokano, T.; Astrobiology 2009, 9, 147. 83. Ruiz, J.; Icarus 2003, 166, 436; Lellouch, E.; Bergh, C.; Sicardy, B.; Ferron, S.; Kauf, H. U.; Astron. Astrophys. 2010, 512, L8. 84. Rampelotto, P. H.; American Geophysical Union Abstracts Book, Foz do Iguaçu, Brasil, 2010; Irwin, L. N.; Schulze-Makuch, D.; Cosmic Biology, Springer: New York, 2011.
Fuente: Quimica Nova, Volumen 35, No. 8, 1619-1627, 2012

Manfenix- Admin
- Mensajes : 3440
Fecha de inscripción : 20/03/2010
Localización : Argentina -

![]()
![]()
![]()
» Es la ley de la vida...
» La vida es...
» La vida interior de una célula
» La Vida Secreta del Caos ♦ Documental BBC
» Diseño Inteligente ♦ El misterio de la vida
» La vida es...
» La vida interior de una célula
» La Vida Secreta del Caos ♦ Documental BBC
» Diseño Inteligente ♦ El misterio de la vida
Página 1 de 1.
Permisos de este foro:
No puedes responder a temas en este foro.